Рецеccивные, доминантные, летальные гены, мутации. III.2.8
До сих пор мы рассматривали генетику человека по типам наследования, теперь мы обратимся к одной очень важной категории генов, а именно к летальным и сублетальным генам. Летальные и сублетальные гены представляют собой класс мутаций с заметно выраженным отрицательным влиянием на развитие. Как мы увидим дальше, мутации в очень многих случаях отягощают и разрушают наследственность человека. Это


отягощение мутациями особенно ярко проявляется под действием летальных генов.
Летальные и сублетальные аллели могут быть доминантными и рецессивными. В случае доминантного леталя ребенок погибает во внутриутробном периоде или в раннем младенческом возрасте. Вполне понятно, что такие гены не могут передаваться по наследству. Их наличие говорит о появлении мутаций. Суммарная частота таких доминантных леталей не так мала. Особенно их количество велико при действии радиации. В генетике мышей и дрозофилы при анализе зависимости частоты мутаций от дозы и при анализе других особенностей мутагенных воздействий широко используются доминантные летали. При наличии сублеталей, иначе называемых полулетальными генами, ребенок умирает в раннем детстве или несколько позднее, но, как правило, до достижения половой зрелости. Примером доминантной сублетальной мутации может служить эпилойя - заболевание, характеризующееся действием целого синдрома отрицательных признаков. Дети, больные эпилойей, имеют патологические разрастания кожи, им свойственна умственная отсталость, эпилепсия, опухоли сердца, почек и других органов. В редких случаях больные эпилойей выживают и вступают в брак со здоровым супругом. Половина детей от такого брака оказывается больными врожденной эпилойей (рис. 131).
Другим ярким примером сублетальной доминантной мутации может послужить заболевание ретинобластома. Этот


доминантный ген вызывает раковую опухоль в глазу, что почти всегда ведет к смерти ребенка в раннем возрасте.
Однако основная масса врожденных уродов падает на рецессивные летали и сублетали. В этом случае рецессивные вредные гены скрываются в генотипе внешне здоровых родителей, однако, когда две таких гетерозиготы вступают в брак, среди их детей около 25% оказываются больными.
На рисунке 132 представлены основные типы скрещиваний в популяциях людей, которые осуществляются при наследовании рецессивных летальных и сублетальных генов. В квадрате 1 представлен тип браков между генетически нормальными людьми. У обоих родителей и у всех их потомков все аллели представлены нормальными генами (аллель +). В квадрате 2 изображен брак между генетически здоровым человеком и гетерозиготой (символ а). В этом случае отрицательный аллель передается половине потомства, оставаясь, однако, при этом в скрытом виде. В квадрате 3 генетически здоровый человек вступил в брак с больным (символ аа), все дети внешне здоровы, но в гетерозиготном состоянии несут сублетальную мутацию а. В квадрате 4 гетерозигота вступает в брак с больным; половина их детей внешне здорова, но несет сублетальную мутацию, другая половина больные. Наконец, квадрат 5 при браке двух больных (символы аа) все потомство будет больным. В качестве врожденных болезней, вызываемых действием рецессивных летальных и сублетальных генов, можно назвать детскую амавротическую идиотию, при которой мозговая ткань дегенерирует, наступает слепота и ребенок гибнет; врожденную патологию кожи с наличием
глубоких кровоточащих трещин - ихтиоз (рис. 133); врожденный детский паралич и целый ряд других врожденных заболеваний.
Многие ранние самопроизвольные выкидыши вызываются выщепляющимися летальными генотипами.
Известны доминантные мутации, в гетерозиготах определяющие какой-либо более или менее нейтральный признак, которые, однако, в гомозиготном состоянии оказывают летальное действие. Так, например, аллель пигментации ксеродермы в гетерозиготном состоянии вызывает сильную веснушчатость. Однако у гомозигот развивается пигментная ксеродерма. Это сублетальное заболевание (рис. 134), при котором действие света вызывает поражение кожи, приводящее затем к злокачественному росту.
Один из аллелей у человека в гетерозиготном состоянии вызывает избыточное содержание холестерина в крови. Однако дети, гомозиготные по этому аллелю, заболевают тяжелой сублетальной аномалией.
Во всех этих случаях мы имеем классическую картину наследования доминантов с рецессивным летальным эффектом, ранее установленную в опытах с дрозофилой, мышами и другими организмами. Например, у мыши хорошо известен аллель A Y , который у гетерозигот вызывает желтую окраску шкурки. Однако все попытки получить чистую расу желтых мышей остались безуспешными. При скрещивании двух желтых мышей в среднем всегда на две желтые мыши рождается одна другой окраски, например черная. Природа этого явления нашла свое объяснение в том, что ген доминантной желтой окраски оказался
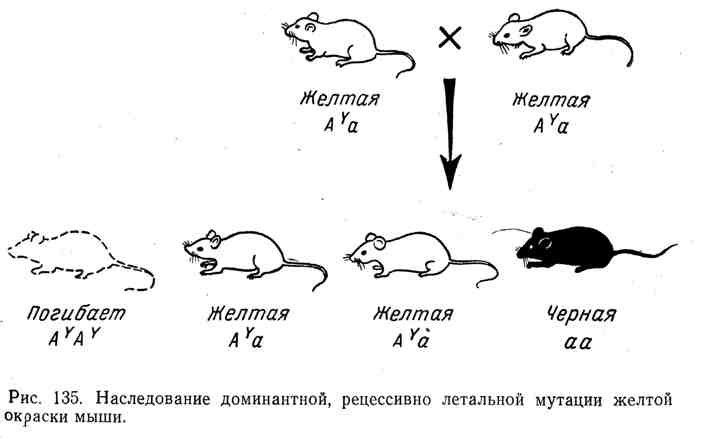
рецессивным леталем. Все гомозиготы ( A Y A Y ) погибали. Желтая мышь всегда гетерозиготна, наряду с аллелем A Y она содержит другой аллель, например а (черной окраски). Ход наследования при скрещивании гетерозигот A Y a и расщепления по формуле 2: 1 изображены на рисунке 135.
— Источник—
Дубинин, Н.П. Горизонты генетики/ Н.П. Дубинин. – М.: Просвещение, 1970.- 560 с.
Post Views: 121
Неодинаковая жизнеспособность зигот разного генотипа может быть связана с доминантными или рецессивными летальными мутациями, возникающими в гаметах родительских особей. Они могут реализовываться на разных стадиях эмбриогенеза или в постэмбриональном периоде. Естественно, что гибель части зигот влияет на характер расщепления.
Влияние на расщепление доминантных генов с рецессивным летальным действием
Доминантные гены с рецессивным летальным действием обладают плейотропным эффектом; с одной стороны, они определяют доминантный характер проявления какого-либо признака в гете-розиготе, с другой - вызывают гибель особей в гомозиготе, т. е. проявляют свое летальное действие как рецессив. Такие гены известны у многих животных - желтая окраска мышей, корот-коногость у кур, линейная чешуйчатость у карпов, платиновая окраска у лис, окраска ширази у овец и многие другие. Они вызывают отклонение от расщепления 3:1, превращая его в расщепление 2:1. Анализ расщеплений с участием доминантных генов с рецессивным летальным действием осложняется тем, что на сравнительно небольших выборках расщепление 3:1 и 2:1 не всегда возможно различить , а статистический метод проверки по критерию χ 2 не позволяет делать выбора из гипотез. Например, если при скрещивании желтых мышей между собой получится расщепление 65 желтых: 32 черных, по методу %2 не отвергаются две гипотезы: 1 - расщепление в опыте соответствует расщеплению 3:1, χ 2 = 3,29, p>0,05; 2 - расщепление в опыте соответствует расщеплению 2:1, χ 2 = 0,17, p>0,05.
Только на больших выборках расщепление 2:1 отличимо от расщепления 3:1 . При суммировании данных по наследованию желтой окраски у мышей, полученных разными исследователями, расщепление при скрещивании желтых мышей между собой было 2386 желтых: 1235 черных - ∑ = 3621, χ 2 = 0,96 при Н 0 - 2:1 (при Н 0 - 3:1 в этом случае χ 2 = 160!).
Очевидно, что разные способы генетической проверки, например постановка анализирующего и возвратного скрещиваний, могут способствовать правильному решению вопроса о наследовании. В рассмотренном выше примере это скрещивания: желтые из F 1 × черные; желтые из F 1 × желтые Р. В первом случае должно быть расщепление 1:1, во втором - снова 2:1.
Сложнее выяснить, что отклонение есть и оно обусловлено разной жизнеспособностью зигот в том случае, когда гибель части зигот осуществляется на постэмбриональной стадии. В связи с этим необходимо вести контроль плодовитости и отхода в потомстве, если есть предположение о наличии летальности. Основанием для предположения о летальности является невозможность получения единообразного потомства при разведении в себе особей определенного фенотипа. Примеры анализа даны в задачах № III.2 и III.3.
Задача № III.2
В потомстве от скрещивания серебристо-соболиных норок между собой в F 1 всегда наблюдается расщепление: появляются серебристо-соболиные и коричневые особи.
В одном опыте в нескольких пометах от скрещивания серебристо-соболиных особей было получено 93 серебристо-соболиных и 43 коричневых щенка при средней величине помета 3,66 щенка. От скрещивания серебристо-соболиных с коричневыми норками получили 39 коричневых и 41 серебристо-соболиных. В скрещиваниях коричневых норок между собой получали только коричневых I потомков. Величина помета в двух последних скрещиваниях составляла 4,9-5,2 щенка.
Объяснить расщепление, определить генотипы серебристо-соболиных и коричневых норок.
Анализ
Поскольку серебристо-соболиные норки при скрещивании между собой и с коричневыми всегда дают расщепление, они очевидно гетерозиготны. Для определения числа генов рассмотрим расщепления в опыте:
H 0 - различия по одному гену, расщепление 3:1, χ 2 = 3,2, p>0,05. Отклонение случайно, гипотеза не отвергается.

Соотношение хорошо соответствует расщеплению по одному гену 1:1, χ 2 = 0,05, p>0,80. Отклонение случайно, гипотеза не отвергается.
Сопоставление данных о плодовитости норок в разных скрещиваниях говорит о частичной гибели зигот при скрещивании серебристо-соболиных норок. Можно предполагать гибель доминантных гомозигот. Тогда генотип серебристо-соболиных норок - Аа, коричневых - аа и расщепление в скрещивании не 3:1, а 2:1 (χ 2 = 0,17, p>0,05). Для окончательного утверждения этой гипотезы необходимо поставить дополнительное скрещивание между серебристо-соболиными особями для увеличения выборки и статистической проверки гипотезы 2:1. В некоторых случаях приходится проводить более сложный анализ.
Задача № III.3
В скрещиваниях платиновых, беломордых и грузинских белых лис с серебристо-черными было установлено, что платиновая, беломордая и грузинская белая окраски, вызывающие общее ослабление пигментации и появление различной пегости, не сцеплены с полом. При скрещивании каждого из этих мутантов с серебристо-черными получали расщепление 2:1, причем серебристо-черная окраска была рецессивной. Следовательно, каждая из них контролируется одним доминантным геном с рецессивным летальным действием. О гибели части мутантов свидетельствуют данные о величине помета: серебристо-черные имеют 4,5 щенка на помет, беломордые 3,5, платиновые и грузинские белые - меньше 3,5.
Для того чтобы установить, затрагивают ли эти мутации один ген или разные, были поставлены скрещивания, результаты которых приведены ниже. Тест на аллелизм в данном случае неприменим, так как мутации доминантные:

Объяснить расщепления, определить генотип всех форм.
Анализ
1. Расщепления в скрещиваниях 1 и 2 соответствуют соотношению 1:1:1:1 (χ 2 = 5,83, p>0,10 для скрещивания 1, χ 2 = 0,55, p>0,90 для скрещивания 2). Они могут быть следствием нескольких причин.
1. Поскольку в скрещиваниях 1 и 2 в расщеплении 4 класса и соотношение 1:1:1:1, можно предположить, что исследуемые формы различаются по двум независимо наследуемым доминантным генам с рецессивным летальным действием, взаимодействующим по типу комплементарности. При этом один из генов представлен двумя разными доминантными аллелями.

2. Расщепление 1:1:1:1 может быть следствием тесного сцепления этих двух генов при отсутствии кроссинговера между ними:

(В решетках Пеннета даны фенотипические радикалы - гены, проявляющие свое действие в фенотипе.)
3. Различия в окраске обусловлены тремя независимо наследуемыми доминантными генами с рецессивным летальным действием. В первом (а) и втором (б) скрещиваниях расщепление происходит по двум разным генам.

4. Расщепление 1:1:1:1 может быть следствием тесного сцепления трех взаимодействующих генов при отсутствии кроссинговера.

* (Порядок расположения генов может быть иным, он дан произвольно. )
5. Расщепление в соотношении 1:1:1:1 может быть результатом различий по одному гену, представленному серией из четырех аллелей, три из которых - доминантные с рецессивным летальным действием, а четвертый - рецессивный:

Для того, чтобы сделать выбор между этими гипотезами, было поставлено скрещивание - белых с серебристо-черными.
Если бы признак контролировался двумя или тремя генами, то при их независимом наследовании можно ожидать появления четырех фенотипических классов:

Аналогичный результат должен быть в скрещиваниях белых, полученных от платиновой или грузинской белой лис, с серебристо-черными, однако вместо беломордой при этом должны были появиться платиновые, в чем легко можно убедиться, написав соответствующие скрещивания.
Полученное в проверочном скрещивании расщепление - появление беломордых и грузинских белых лис - можно объяснить либо взаимодействием двух (или трех) тесно сцепленных генов, либо взаимодействием трех аллелей при моногенном различии между исходными формами.

Резкое снижение жизнеспособности белых лис, отмеченное в опыте, говорит в пользу действия аллельных мутаций, так как в этом случае белые лисы по генотипу являются компаундами по двум доминантным мутациям одного гена, причем обе - с рецессивным летальным действием. Трудно ожидать снижения жизнеспособности при взаимодействии двух разных генов (комплементарность). Поэтому сделали вывод, что белая, беломордая, грузинская белая, платиновая и серебристо-черная окраски у лис контролируются серией аллелей одного гена, три из которых - доминантные с рецессивным летальным действием. Генотипы лис: белая A 1 / A 2 , A 1 / A 3 , A 2 / A 3 ; беломордая A 1 / а; платиновая A 3 / а; грузинская белая A 2 / а; серебристо-черная а / а (по Беляеву и др., 1973).
Следует подчеркнуть, что при множественном аллелизме в случае гетерозиготности исходных форм максимальное число фенотипических классов в расщеплении может быть не 3, а 4, как в случае, описанном выше. В популяции же число возможных генотипов при множественном аллелизме многократно возрастает; оно может быть определено по формуле: 1 / 2n (n+1), где n - число аллелей. Например, если имеется 7 аллелей какого-либо локуса, число возможных генотипов в популяции будет равно 28: 1 / 2 ×7×8 = 28.
Влияние рецессивных летальных мутаций на расщепление
Рецессивные аутосомные и сцепленные с полом летали, вызывая гибель гомозигот в скрещиваниях гетерозигот по летали, могут влиять на расщепление по генам, сцепленным с леталькк При этом соотношение потомков в расщеплении зависит от расстояния между изучаемым геном и леталью, а также от типа гетерозиготы - в цис- или транс-положении гены вводятся в гетерозиготу. Для выявления летали обычно ставятся разные проверочные скрещивания. Примером анализа летали служит задача № III.4.
Задача № III.4
В линии дрозофилы из линии № 100, содержащей инверсии, половина самок имели серую, половина - желтую окраску тела, причем желтые самки оказались стерильными. Все самцы этой линии имели желтую окраску. Соотношение самок и самцов отличалось от нормального, оно больше похоже на расщепление 2♀♀:1♂♂. Было высказано предположение, что недостаток самцов в линии № 100, вероятно, связан с наличием летали в Х-хромосоме серых самок, которые, очевидно, гетерозиготны - в линии происходит расщепление на серых и желтых самок. Для установления генетической структуры этой линии и проверки предположения о наличии летали в Х-хромосоме самок были поставлены скрещивания, результаты которых представлены ниже.
Реципрокные скрещивания

Серые самки из F 1 были индивидуально скрещены с желтыми самцами из линии № 100.

Анализ
На основании проведенного анализа запишем схемы всех скрещиваний.

Предложенные гипотезы достаточно хорошо объясняют все результаты. Однако остается невыясненным вопрос о причине стерильности желтых самок в линии № 100. Предлагаем читателю обдумать этот вопрос и предложить какую-либо гипотезу для его объяснения.
У растений многие рецессивные мутации связаны с недостатком или отсутствием хлорофилла, что приводит либо к снижению жизнеспособности растений, либо к их гибели на разных этапах развития. Это служит причиной отклонений в расщеплении, а также вызывает необходимость учета расщеплений не только на проростках, но и на более поздних стадиях развития для определения доли гибели растений и характера наследования признака. Так, у кукурузы гомозиготы по гену wd (white deficiency) имеют белые проростки (расщепление на проростках 3 / 4 зеленых: 1 / 4 белых). Однако спустя 1-3 недели происходит отмирание всех белых растений после использования запасов питательных веществ семени и в наиболее поздних стадиях развития растений расщепление исчезает - 3 зеленых: 0 белых. Аналогичные мутации известны у гороха, ячменя, ржи, пшеницы и др.
Другие мутации вызывают гибель лишь части особей на определенном этапе развития, что приводит к уменьшению доли рецессивов в расщеплении и изменению соотношения фенотипов - 4:1, 5:1 и т. п. Эти соотношения варьируют, поскольку, как правило, жизнеспособность таких мутантов в значительной степени зависит от условий.
У человека пониженная жизнеспособность и летальность вследствие действия рецессивных мутаций проявляется в разные периоды эмбриогенеза и на разных этапах развития. Причины снижения жизнеспособности и летального эффекта могут быть связаны как с генными мутациями, так и с хромосомными аномалиями. Цитогенетический анализ абортивных зародышей позволяет установить причину гибели многих из них. В среднем из-за аберраций хромосом на всех сроках беременности происходит более 42% спонтанных абортов: значительная часть новорожденных с хромосомными аберрациями погибает в течение первого и последующих лет жизни.
Среди генных летальных мутаций, приводящих к внутриутробной гибели или смерти в младенческом возрасте, можно назвать рецессивные мутации, вызывающие талассемию, серповидноклеточную анемию, муковисцедоз, врожденный ихтиоз, анэнцефалию (отсутствие мозга), фенилкетонурию и др.
Для изучения летальных или понижающих жизнеспособность мутаций у человека широко используются цитогенетический и биохимический методы анализа, изучение структуры и активности ферментов в норме и при патологии, а также у гетерозиготных носителей; хроматография, разные типы электрофореза.
Летальные гены – мутационные гены, вызывающие гибель особи до достижения ею половозрелого возраста. Бывают доминантными, рецессивными, сцепленные с полом. Обычно проявляют своё действие в гомозиготном состоянии, в гетерозиготном состоянии понижают жизнеспособность. Пенетрантность – способность гена проявиться фенотипически, выражается в % и бывает полный и неполный. Полный – у всех особей популяции, имеющих данный ген, он проявляется в виде признака. Неполный – у некоторых особей ген имеется, но внешне себя не проявляет. Экспрессивность – степень проявления признака, т.е. один и тот же признак у разных особей выражен с разной интенсивностью.
При наличии разных летальных генов организмы гибнут на различных стадиях развития. Как правило, летальное действие таких генов рецессивно, т.е. проявляется лишь тогда, когда они находятся в гомозиготном состоянии. При возникновении мутаций с доминирующим летальным действием организм погибает, не давая потомства.
Однако отмечены случаи, когда летальные гены, вызывая в гетерозиготном состоянии видимые изменения, становятся даже полезными с хозяйственной точки зрения. Так, среди овец каракульской породы встречаются животные с красивой серебристо-серой окраской шкурки, оцениваемой дороже, чем обычный черный каракуль, При скрещивании серых овец с серыми баранами оказалось, что они всегда гетерозиготны. При изучении причин этого явления было установлено, что среди полученных в результате такого скрещивания серых ягнят примерно 1/3, или около 25% всего приплода, заболевала хроническим тимпанитом и погибала. Причиной заболевания служили нарушения в деятельности парасимпатической нервной системы. Когда же скрещивали серых баранов с черными овцами или серых овец с черными баранами, в потомстве рождалось 50% серых и 50% черных ягнят, причем серые ягнята не заболевали. Оказалось, что в гомозиготном состоянии ген, вызывавший развитие серой окраски, обладал рецессивным летальным действием.
У лисиц встречается ценная окраска меха - платиновая, обусловленная доминантным геном, вызывающим в гомозиготном состоянии гибель зародышей на ранней стадии развития. В таких случаях соотношение фенотипов во втором поколении меняется, так как из трех доминантных форм одна (гомозиготная по летальному гену) погибает, в результате чего соотношение фенотипов становится 2:1.
Аналогичное расщепление отмечено и у зеркального карпа, у которого наблюдалось наследование недоразвития чешуи: она сохранилась только на средней линии тела, остальная часть его голая, поэтому его называют линейным. При скрещивании между собой линейных карпов потомство всегда расщепляется в соотношении 2 линейных: 1 с нормальной чешуей, так как зародыши, гомозиготные по гену, вызывающему недоразвитие чешуи, погибают на ранних стадиях развития.
У кур, гомозиготных по аллелю, вызывающему «курчавость» перьев, неполное развитие перьев влечет за собой несколько фенотипических эффектов. У таких кур теплоизоляция недостаточна, и они страдают от охлаждения. Для компенсации потери тепла у них появляется ряд структурных и физиологических адаптаций, но эти адаптации малоэффектны и среди таких кур высока смертность.
Воздействие летального гена ясно видно на примере наследования окраски шерсти у мышей. У диких мышей шерсть обычно серая, типа агути; но у некоторых мышей шерсть желтая. При скрещиваниях между желтыми мышами в потомстве получаются как желтые мыши, так и агути в отношении 2:1. Единственное возможное объяснение таких результатов состоит в том, что желтая окраска шерсти доминирует над агути и что все желтые мыши гетерозиготны. Атипичное менделевское отношение объясняется гибелью гомозиготных желтых мышей до рождения. При вскрытии беременных желтых мышей, скрещенных с желтыми же мышами, в их матках были обнаружены мертвые желтые мышата. Если же скрещивались желтые мыши и агути, то в матках беременных самок не оказывалось мертвых желтых мышат, поскольку при таком скрещивании не может быть потомства, гомозиготного по гену желтой шерсти.
Изменение расщепления по фенотипу в отношении 3:1 во втором поколении моногибридного скрещивания бывает связано с разной жизнеспособностью зигот Ф2. Разная жизнеспособность зигот может быть обусловлена наличием летальных генов. Летальным называется ген, вызывающий нарушения в развитии организма, что приводит его к гибели или уродству.
Изучение врожденных аномалий показало, что при разных летальных генах гибель особей бывает различной и может происходить на разных стадиях развития.
По классификации, предложенной Розенбауэром (1969), гены, вызывающие гибель 100 % особей до достижения ими пЪловой зрелости, называются летальными, более 50 % --сублетальными (полулетальными) и менее 50 % -- субвитальными. Однако следует отметить, что разделение это до некоторой степени условно и иногда не имеет четких границ. Примером может служить сцепленная с полом голость у кур. Почти половина голых цыплят гибнет в последние 2--3 дня инкубации. Из числа вылупившихся около половины цыплят гибнет до 6-недельного возраста, если их выращивают при температуре 32--35 "С. Но если температура в брудерах будет повышена на 5,5 °С, то погибнет значительно меньше голых цыплят. В 4--5 мес у голых цыплят вырастает редкое оперение и они уже в состоянии переносить довольно низкие температуры.
В естественных условиях эта мутация, по-видимому, будет летальной и приведет к 100%-ной гибели птиц. Приведенный пример показывает, что характер проявления полулетального гена в значительной мере может зависеть от условий среды.
Летальные гены могут быть доминантными и рецессивными. В числе первых летальных факторов был открыт аллель, обусловливающий желтую окраску мышей. Ген желтой окраски является доминантным (Y). Его рецессивный аллель (у) в гомозиготном состоянии обусловливает появление черной окраски. Скрещивание желтых мышей между собой давало в потомстве две части особей желтых и одну часть черных, т. е. получалось расщепление 2:1, а не 3:1, как следовало из правила Менделя. Оказалось, что все взрослые мыши гетерозиготны (Yy). При скрещивании между собой они должны были дать одну часть гомозиготного потомства по желтой окраске (IT), но оно погибает еще в эмбриональном периоде, две части гетерозигот (Yy) будут желтыми и одна часть гомозигот по рецессивному признаку (уу) будет черной. Схема скрещивания выглядит так:
Таким же образом наследуется серая окраска шерсти у каракульских овец (сокольских, малич и др.), платиновая окраска у лисиц, распределение чешуи у линейных карпов и т. д.
Летальные гены в большинстве случаев рецессивны и поэтому долгое время могут находиться в скрытом состоянии.
Совершенно здоровое и нормальное по фенотипу животное может быть носителем летального гена, действие которого обнаруживается только при переходе в гомозиготное состояние. В гомозиготное состояние летальные гены чаще всего переходят при родственном спаривании. В практике животноводства при разведении лошадей был случай гибели 25 жеребят на 2--4-й день после рождения от уродства прямой кишки -- отсутствия заднепроходного отверстия (Atresia ani). Оказалось, что все жеребцы и кобылы, от которых рождались такие ненормальные жеребята, происходили от одного жеребца. Он был гетерозиготен по летальному гену (LI). Вначале этот жеребец при скрещивании с нормальными кобылами (LL) дал потомство, по фенотипу нормальное, но по генотипу половина потомков была благополучной (LL), a половина -- гетерозиготной (LI), несущей рецессивный задаток (0 летального гена. При родственном спаривании гетерозиготных животных (Ы х Ы) появилась часть жеребят, гомозиготных по летальному гену (II), с уродством прямой кишки. Они все погибли.
У сельскохозяйственных животных известны десятки аномалий, возникновение которых связано с рецессивными или доминантными мутациями генов. Эти аномалии встречаются в отдельных популяциях с разной частотой, что зависит от скорости мутационного процесса, системы разведения животных и др. Знание конкретных форм врожденных аномалий у животных каждого вида, а также частоты их проявления в отдельных породах необходимо ветеринарным специалистам для селекционной профилактики распространения генетической патологии.
Аномалии у крупного рогатого скота. Биологические особенности данного вида животных - малошюдие и относительная позднеспелость. Корова обычно приносит одного теленка, кото-, рый достигает половой и физиологической зрелости только к 1,5 года, так что период между первым отелом матери и дочери в среднем составляет 5 лет. Вследствие этого появление в стаде аномального приплода может существенно снижать уровень воспроизводства и интенсивность племенного отбора поголовья. У крупного рогатого скота изучен широкий спектр врожденных шомалий, детерминированных летальными, полулетальными и субвитальными генами. 46 аномалий включено в Международный список летальных дефектов под шифром А (табл. 43). Относительная частота отдельных типов аномалий в каждой породе или популяции может быть различной. В костромской породе, по нашим данным, наиболее часто регистрируется генетическая аномалия головы - укорочение челюсти (табл. 44), в ярославской породе - синдактилия, в холмогорской - контрактуры мышц, в черно-пестрой - пупочные грыжи. У крупного рогатого скота в Германии наиболее часто (21 %) встречались аномалии центральной нервной системы.
Второе место по частоте регистрации (14 %) занимала комплексная аномалия - сочетания пупочных грыж с расщеплением брюха и плода в целом. Частота аномалий, или процентное отношение аномального приплода к общему его числу, в пределах конкретных популяций также может быть самым различным и по средним оценкам не превышать 1 %. Однако этот показатель зависит от полноты и точности регистрации аномалий. Так, в Германии после организации четкого учета сделали вывод, что частота аномалий возросла в несколько раз. Вопрос в другом: все ли аномалии поддаются визуальному наблюдению? Очевидно, не все. Так, в костромской породе средняя частота всех форм аномалий за 12-летний период составила 1,15 %. Частота же общей смертности приплода (абортированные, мертворожденные, аномальные, павшие без видимых дефектов телята) в этом хозяйстве составила 10,2 %. Определенная доля этой смертности также связана с генными мутациями, вызывающими не морфологические дефекты, а нарушение обмена и другие аномалии, выявление которых возможно только специальными методами.
Особую роль в распространении генетических аномалий как у крупного рогатого скота, так и у животных других видов могут сыграть производители. От каждого производителя при искусственном осеменении в год можно получить сотни и тысячи потомков. Так, от одного быка за рубежом получили 100 тыс. телят. Если такой производитель окажется носителем генной мутации, то она быстро распространится в породе. Вот некоторые примеры из многочисленных фактов, описанных в литературе. В результате интенсивного использования быка Принца Адольфа, завезенного в Швецию, и последующего стихийного инбридинга на него частота бесшерстности в отдельных шведских стадах была свыше 5 %. Такая же ситуация сложилась в Швеции после импорта быка Галлуса, который оказался гетерозиготным носителем гена, обусловливающего отсутствие конечностей.
В потомстве отдельных быков черно-пестрой породы и Шароле в США и Германии регистрировали случаи рождения карликовых телят с частотой соответственно 23,3 и 22,2 %. В бывш. Чехословакии при обследовании потомков 166 быков-производителей установлено, что 43 из них были носителями летальных генов. У одного быка - носителя доминантной аномалии «заячья губа»-дефект проявился среди 44 % бычков и 71 % телочек из его потомства.
В костромской породе нами проанализировано распространение укорочения нижней челюсти и мопсовидности через быка Бурхана, который сам имел в потомстве дефектных телят; его сыновья, внуки, правнуки, женские потомки также давали аномальный приплод (рис. 57). Большинство телят получено от родственного спаривания и спаривания родителей с нормальным фенотипом при наличии общего предка. Следовательно, можно сделать вывод о рецессивном типе наследования данной аномалии. Из рисунка видно, что наибольшее количество дефектных телят зарегистрировано в потомстве быка Жетона 3501 (внука быка Бурхана) при использовании его в товарном хозяйстве, где часть коров имела в генотипе такой же рецессивный ген.
Аномалии у свиней. В Международный список летальных дефектов свиней включено 18 генетических аномалий. Основная часть их обусловлена аутосомными рецессивными генами (табл. 45). Генетические аномалии могут занимать значительное место в патологии свиней. Рассмотрим несколько таких примеров. В Испании при исследовании 23 449 поросят из 2399 пометов, полученных от хряков пород дюрок, йоркшир, гемпшир и белый честер, выявили соответственно 6,21; 6,02; 9,66; 2^62 % анрмаль-ных пометов.
Согласно данным Оливье (1979), у свиней описано 7 генетических аномалий кожного покрова, 17 - скелета, 3 - глаз, 13 - нервно-мышечных, 6 - крови, 6 - гормонально-обменных, 5 - пищеварительной системы, 9 - мочеполовой. Основными аномалиями были крипторхизм, грыжи, псевдогермафродитизм и др. Автор исследования считает, что эти аномалии - результат действия одного гена на разных этапах формирования эмбриона.
В Дании для выяснения характера и частоты возникновения врожденных аномалий в течение двух лет исследовали 6669 павших поросят из 2936 пометов. Различные аномалии выявлены у 1,4 % поросят к числу родившихся, или 6,2 % к числу павших до отъема особей. При патологоанатомическом исследовании у 25,9 % аномальных поросят были обнаружены недоразвитие клапанов, незакрытие анального отверстия, субаортальные стенозы, эктопия сердца и другие дефекты сердечно-сосудистой системы. У 23,4 % поросят найдены различные нарушения развития двигательной системы. Аномалии центральной нервной системы выявлены у 5,9 % поросят, среди них раздвоенный мозг и водянка мозга. Заращение прямой кишки, тонких кишок или их неполное развитие обнаружены у 30 % поросят, а различные грыжи и асцит - у 6,8 %. «Заячья губа», расщепление нёба, риноцефалит и другие аномалии лицевой части головы найдены у 6,1 %; гермафродитизм, незаращение мочеточника, водянка почек и уретры-у 1,7 % поросят. Указанные аномалии проявлялись в потомстве отдельных производителей при инбридинге, что указывает на наследственный характер их возникновения.
Весьма убедительные доказательства наследственной природы крипторхизма у свиней получили Фридин и Ньюман. По их данным, в Канаде одно-, двухсторонний крипторхизм наблюдается ежегодно у 1-2 % всех хрячков, поступающих на рынок. Авторы скрещивали крипторхов с их матерями и полными сестрами. Потомков от таких скрещиваний спаривали между собой. В результате такого отбора и подбора частота крипторхизма у экспериментальных животных йоркширской породы возросла в среднем до 42,9 % и особенно при использовании двух производителей. При обследовании поросят в США за один год было обнаружено около 400 тыс. животных с мошоночной грыжей.
Наблюдения показывают, что причиной нарушения плодовитости у хряков нередко бывает гипоплазия семенников. Частота этой аномалии, по данным исследователей из Германии, составила 19,6 %. 30 таких хряков были оставлены для воспроизводст-
Ва, каждым из них было покрыто от 4 до 40 маток (всего 439 голов), но лишь четыре из них дали потомство. Анализ показал, что патологические формы спермиев у этих хряков составляют 80-100 %. Все 30 аномальных животных имели общих предков, что указывает на наследственный характер гипоплазии семенников и дефектов спермиогенеза.
Наличие кратерных сосков у свиней - один из серьезных дефектов, поскольку поросята не получают от них молока. По данным Баварского института животноводства (Германия), частота этой аномалии у немецких ландрасов составила 6,6 %. Как отмечают П. Н. Кудрявцев и др. (МВА), количество свиней с бездействующими кратерными сосками в последние годы увеличилось. Число таких сосков колеблется от 1 до 8. Поросята, которым достаются кратерные соски, погибают.
Кратерность - признак, обусловленный одним аутосомным рецессивным геном. Экспериментально это проверили П. Н. Кудрявцев с сотр. Предварительно выделив хряков и свинок-носителей аномалий (кчкч), нормальных, но гетерозиготных особей (Кчкч), дающих в приплоде поросят с кратерностью, и нормальных гомозиготных свиней (КчКч), авторы провели скрещивания между этими группами животных. В первом варианте скрещивали 27 нормальных гомозиготных маток с 15 хряками. Все 258 потомков были нормальными. Во втором варианте, где один из родителей был гомозиготным (КчКч), а другой - гетерозиготным (Кчкч), все поросята также были нормальными. В третьем варианте скрещивали 13 гетерозиготных хряков с 16 гетерозиготными матками. Из 168 родившихся поросят 39 (23,2 %) имели кратерные соски. И наконец, в четвертом варианте один из родителей был гомозиготным, а другой - гетерозиготным. От них было получено 170 поросят, из них 86 (50,5 %) оказались с нормальными сосками и 84 (49,5 %) - с кратерными. Результаты этого опыта доказывают рецессивный тип наследования кратерности сосков у свиней.
Аномалии у овец. У овец описано около 90 врожденных аномалий. По данным Денниса и Лейпольда, большинство известных генетических дефектов у овец обусловлено моногенным аутосомным рецессивным типом наследования (табл. 46). Наиболее часто у этого вида животных встречаются черепно-лицевые дефекты, особенно агнатия, а также искривление передних конечностей, микроагнатия, гермафродитизм, крипторхизм, гипос-падиаз, прогнатия, атрезия ануса, микротия, энтропия, тортико-лис, полителия, артрогрипоз. Анализ показал, что 55,4 % дефектов относились к мышечно-скелетной системе, 12,7 - к пищеварительной, 9,7 - к сердечно-сосудистой, 7,1 - к урогени-тальной, 6 - к центральной нервной системе, 3,5 - к аномалии связок, 3,2 - к брюшной, 1,5 % - к эндокринной системе. Хотя частота отдельных дефектов и невысока, однако кумулятивный вклад всех аномалий может нанести ущерб хозяйствам. В Новой Зеландии, в стране развитого овцеводства, частота летальных дефектов составила около 1 % павших ягнят. В США летальные дефекты в первые 2 нед жизни наблюдались у 11,4 % ягнят.
Средняя эмбриональная смертность у овец равна 20 %. Это указывает на то, что в этот период могут проявлять свое действие многие неидентифицируемые летальные гены.
Отдельные генетические аномалии у овец могут быть широко распространены. Так, в Болгарии в стадах мериносовых овец наблюдали высокую смертность ягнят в ранний постнатальный период. Она наступала в результате того, что ягнята не получили молока из-за дефектов вымени их матерей: от гипоплазии с небольшими остатками железистой ткани до полного отсутствия ее. Частота этой наследственной аномалии в разных стадах колебалась от 6 до 40 %.
У овец часто наблюдали крипторхизм, который сочетался с таким вторичным половым признаком, как комолость. Комолые бараны имеют низкую плодовитость. Путем жесткого племенного отбора удалось создать тип комолых баранов с нормальной плодовитостью, однако и среди них встречаются аномальные особи.
Аномалии у птиц. Птицы, прежде всего куры, наиболее изучены в отношении генетики аномалий. В Международный список летальных дефектов включено 45 аномалий у кур, 6 у индеек и 3 у уток. Наиболее часто встречаются аномалии клюва (клюв попугая, перекрещивающийся клюв). Частота их, по данным Уильяма и др., составляет 1,1 % потерь при инкубации яиц кур пород белый леггорн и род-айланд. Аномалии клюва часто встречаются и у уток.
Макс Джиббон и Шейкелферд описали аномалию при скрещивании белых леггорнов с батеркампами и бентамками и последующем разведении «в себе» - полидактилию. Кроме нее у цыплят наблюдали синдактилию и оперенные ноги. Частота синдрома составляла 16,8 %. При скрещивании аномального петуха F2 с нормальными по фенотипу курами наблюдали расщепление - половина нормальных и половина аномальных особей (1:1). Установлено, что данный комплекс признаков контролируется одним аутосомным геном, обладающим полулетальным эффектом, так как эмбриональная и постэмбриональная выживаемость аномальных цыплят очень низка.
Аномалии у лошадей. Из наследственных аномалии у лошадей 10 включены в Международный список летальных дефектов. Среди них 3 аномалии скелета, 2 - воспроизводительной системы, 2 - почек и мышц, по одной аномалии кишечника, нервной системы, органов зрения.
У лошадей тяжеловозных пород чаще встречается атрезия ободочной кишки. Распространение этой аномалии было отмечено в потомстве жеребца Суперба першеронской породы. Описана она также у жеребят чистокровной верховой породы остф-ризского происхождения. У тяжеловозных пород зарегистрирован несовершенный эпителиогенез. У жеребят ольденбургской породы в Германии обнаружена атаксия, названная ольденбургской. Распространилась она в линии 9. Одна из наиболее часто регистрируемых аномалий у лошадей - пупочная грыжа. Встречается она у легких и тяжеловозных пород.
У лошадей известны некоторые другие генетические и наслед-ственно-средовые аномалии. Так, в США у лошадей нескольких пород наблюдается появление жеребят со своеобразной белой пятнистостью, названной «оверо». При скрещивании лошадей типа «оверо» рождаются жеребята с розовой кожей, у которых наблюдают гипоплазию кишечного тракта и изоэритролиз, а также колики, приводящие к гибели.
В Англии у лошадей дшстокровной верховой породы зарегистрированы и исследованы животные с нарушением координации движений - «болезнь вихляния». Установлена генетическая предрасположенность к этой аномалии.
Доказана наследуемость дерматозов конечностей у лошадей. Пороками с наследственной предрасположенностью являются нередко встречающееся хроническое деформирующее воспаление скакательного сустава - шпат, так называемая «костыльная нога» у жеребят, а также хроническое асептическое воспаление венечного блока копыт, наблюдаемое в основном у скаковых и беговых лошадей.





 (1921) Подавление кронштадтского восстания")



- Лимонное печенье из песочного теста Как приготовить лимонное печенье из песочного теста
- Салат ералаш с говядиной рецепт
- Горбуша, запеченная в духовке с картошкой
- Как приготовить хворост в домашних условиях: вкусные и легкие рецепты
- Домашняя бастурма — лучшие рецепты
- Как обустроить рабочий стол по Фен Шуй для денег
- Заговоры против соперницы вернут спокойствие в семью
- Конспект по обучению грамоте в подготовительной группе «Космическое путешествие
- Чиновник Сергей Рыбаков: «Время – то, что мы в него вмещаем
- Боли в левом боку со стороны спины
- На автомобиле - от государства
- Финансы в экономике. Банковская система. Финансы в экономике Презентация обществознание 11 кл финансы в экономике
- Презентация на тему финансы в экономике
- Происхождение и история аварского народа
- Медицинские приборы для лечения суставов в домашних условиях Бытовой ультразвуковой прибор физиотерапии для лечения суставов
- Территориальные единичные расценки
- Кронштадтское восстание («мятеж») (1921) Подавление кронштадтского восстания
- Даосская система. Л. БингСекреты любви. Даосская практика для женщин и мужчин. Система «Вселенское Дао»
- Цигун: китайская практика укрепления организма
- Оед общество евангелизации детей